 Danh mục cây trồng
Danh mục cây trồng Côn trùng hại cây trồng
Côn trùng hại cây trồng Bệnh hại cây trồng
Bệnh hại cây trồng Dinh dưỡng cây trồng
Dinh dưỡng cây trồng Bài chuyên mục
Bài chuyên mục Liên hệ
Liên hệCơ chế hấp thu Ion của tế bào rễ và sự chuyển vận gần

1. Đặc tính hấp thu ion của cây trồng
Sự hấp thu dinh dưỡng của cây có tính chọn lọc. Dung dịch ở bên ngoài và bên trong tế bào được ngăn cách bởi 2 màng: màng tế bào chất và màng không bào. Sự hấp thu ion của thực vật thượng đẳng có những đặc tính sau:
- Tính chọn lọc. Có dưỡng chất khoáng nầy thì được ưu tiên hấp thu, trong khi chất khác thì bị ngăn cản hay loại trừ.
- Có sự tích lũy dưỡng chất. Biểu hiện nồng độ của dưỡng chất khoáng trong tế bào chất cao hơn dung dịch bên ngoài.
- Theo đặc tính di truyền. Có sự khác biệt giữa các loại cây trồng về đặc tính hấp thu ion.
2. Đường dẫn các chất tan từ bên ngoài vào rễ
2.1. Vận chuyển qua khoảng trống tự do
Các chất tan từ dung dịch bên ngoài đi vào bên trong cây qua vách tế bào và những khoảng trống giữa các tế bào của phần vỏ rễ. Sự di chuyển của các chất tan có trọng khối phân tử thấp (như các ion, acid hữu cơ và các amino acid) bằng cách khuyếch tán không bị hạn chế bởi mặt ngoài của rễ, nghĩa là tế bào biểu bì rễ (Hình 1).
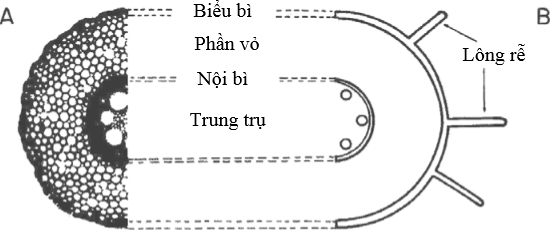
Hình 1 Phẩu diện của rễ bắp
Các khoảng trống tự do chiếm khoảng 10% tổng thể tích rễ non. Sự hiện diện của khoảng tự do này cho thấy rằng các tế bào rễ ở phần vỏ có khả năng hấp thu các chất tan trực tiếp từ dung dịch bên ngoài. Sự khuyếch tán chất tan vào trong khoảng tự do phụ thụộc nhiều yếu tố như nồng độ chất tan và sự hình thành lông rễ.
Cấu tạo vách tế bào sơ lập gồm cellulose, hemicellulose (gồm có pectin) và glycoprotein nên có nhiều lỗ có kích thước khác nhau. Kích thước của các ion như K+ và Ca2+ nhỏ so với kích thước các lỗ (Bảng 1).
Bảng 1 Đường kính lỗ ở vách tế bào và kích thước các ion
|
Đường kính, nm |
|
|
Vách tế bào biểu bì rễ bắp |
500-3.000 |
|
Vách tế bào vỏ rễ bắp |
100-200 |
|
Khoảng trống trong vách tế bào |
<5,0 |
|
Đường sucrose |
1,0 |
|
Các ions thủy hóa: |
|
|
K+ |
0,66 |
|
Ca2+ |
0,82 |
Chất tan có khối lượng phân tử cao (như chelate kim loại, fulvic acid và độc chất) hoặc virút và mầm bệnh bị hạn chế hay ngăn chặn không cho vào bên trong khoảng trống tự do tế bào rễ nhờ kích thước lỗ. Hệ thống này được cấu tạo bao gồm một tỷ lệ của pectin chứa các acid polygalacturonic nên trong khoảng trống tự do có các nhóm carboxylic (R-COO-) hoạt động như chất trao đổi cation. Vì vậy các cation tích tụ lại ở những khoảng trống, trong khi đó các anion bị đẩy ra (Hình 2).

Hình 2 Khoảng trống tự do
Các cation hóa trị hai như Ca2+ ưu tiên liên kết ở các vị trí trao đổi cation. Mỗi loại cây trồng có số lượng nhóm carboxylic khác nhau nên cũng khác nhau về khả năng đổi cation (CEC) ở vách tế bào (Bảng 2). Có tương quan thuận giữa CEC và tỉ số Ca2+/K+ chứa trong khoảng trống tự do. Thường thì CEC ở những loài cây hai lá mầm cao hơn CEC ở những loài cây một lá mầm.
Bảng 2 Khả năng trao đổi cation (CEC) của rễ ở nhiều loài cây khác nhau (Keller và Deuel, 1957)
|
Loài cây |
CEC (meq/100g trọng lượng khô) |
|
Lúa mì |
23 |
|
Bắp |
29 |
|
Đậu |
54 |
|
Cà chua |
62 |
2.2. Vận chuyển qua tế bào chất và không bào
Sự hấp thu chọn lọc các cation và anion xảy ra ở vị trí của màng tế bào. Màng tế bào chất giống như rào cản, ngăn chặn sự khuyếch tán các chất tan vào trong tế bào chất hoặc khuyếch tán chất tan từ tế bào chất đi ra vách tế bào vào dung dịch bên ngoài. Một rào cản khác là màng không bào (tonoplast), khi tế bào đã trưởng thành thì không bào chiếm trên 90% thể tích tế bào và chính là nơi tích lũy các ion (Hình 3).
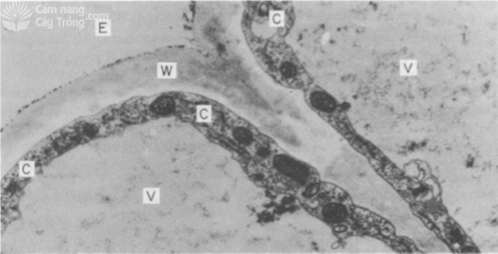
Hình 3 Phẩu diện tế bào ngoại bì rễ bắp. V. Không bào; C. Tế bào chất; W. Vách tế bào; E. Dung dịch bên ngoài
Màng tế bào chất và màng không bào là màng sinh học có liên quan trực tiếp đến sự hấp thu chất tan và vận chuyển chất tan trong rễ. Qua các thí nghiệm về sự
hấp thu cation K+ và Ca2+, người ta đã chứng minh được chức năng của màng tế bào chất và màng không bào như là rào ngăn cản có hiệu quả đối với sự khuyếch tán và sự trao đổi của các ion. Sự vận chuyển chất tan vào các ty thể và lục lạp phải do màng tế bào qui định. Khả năng vận chuyển chất tan qua màng có liên quan chặt tới thành phần hóa học và cấu trúc phân tử của màng.
3. Cấu trúc và thành phần của màng tế bào
Có người cho rằng màng tế bào có những lỗ với kích thước nhất định và sự thẩm thấu của các phân tử qua các lổ trên màng giảm khi đường kính lỗ gia tăng. Nhiều nghiên cứu đã chứng minh một số chất tan đi qua màng nhanh hơn nhiều so với dự đoán dựa trên lý thuyết cơ bản về kích cở của phân tử. Các phân tử này có tính hoà tan cao trong dung môi hữu cơ do có thành phần thu hút mở. Vì vậy, người ta kết luận rằng các lỗ trên màng cũng có thành phần tương tự nên làm cho một số phân tử lớn thu hút mở thấm qua màng nhanh hơn.
Theo nghiên cứu của Branton (1969) và Singer (1972) cho thấy rằng mô hình màng lỏng bao gồm các thể protein và hai lớp phospholipid có những vùng ở đầu tích điện, hút nước (amino phosphate và carboxyl) hướng tới bề mặt màng. Các protein liên kết nhau (protein ngoại sinh) bởi liên kết tĩnh điện hướng tới bề mặt như là enzyme liên kết màng. Các protein khác được kết hợp vào trong màng ở bên trong hay đi ngang qua màng (protein nội sinh) hình thành các “đường rãnh protein” hay còn gọi là protein vận chuyển. Các đường rãnh protein được xem như là những lỗ hút nước làm cho các chất tan như các ion được vận chuyển qua (Hình 4).
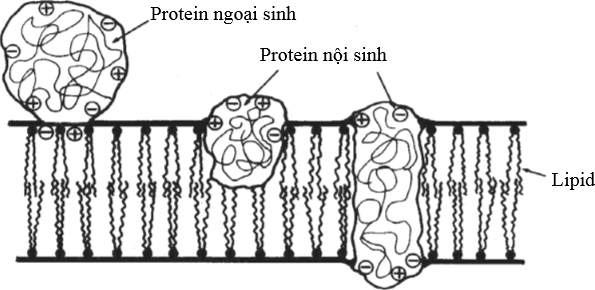
Hình 4 Mô hình màng sinh học
4. Sự vận chuyển chất tan ngang qua màng sinh học
4.1. Sự vận chuyển nhờ chất mang
 Màng tế bào là rào cản hiệu quả để các ion và các phân tử không tích điện đi qua. Hay nói cách khác, màng là nơi hấp thu có chọn lọc, khác với sự vận chuyển dựa vào sự chênh lệch về nồng độ. Tính chọn lọc và sự tích lũy các chất tan ở màng nhờ vào chất mang, chất mang nầy sẽ liên kết với các ion như kali và các phân tử không tích điện và vận chuyển chúng đi qua màng (Hình 5A). Sự vận chuyển nầy nhờ vào năng lượng để kích hoạt chất mang. Năng lượng sử dụng là những phosphate giàu năng lượng dưới dạng ATP được trình bày ở Hình 5B. ATP được tái tạo trở lại từ ADP + Pi qua hô hấp.
Màng tế bào là rào cản hiệu quả để các ion và các phân tử không tích điện đi qua. Hay nói cách khác, màng là nơi hấp thu có chọn lọc, khác với sự vận chuyển dựa vào sự chênh lệch về nồng độ. Tính chọn lọc và sự tích lũy các chất tan ở màng nhờ vào chất mang, chất mang nầy sẽ liên kết với các ion như kali và các phân tử không tích điện và vận chuyển chúng đi qua màng (Hình 5A). Sự vận chuyển nầy nhờ vào năng lượng để kích hoạt chất mang. Năng lượng sử dụng là những phosphate giàu năng lượng dưới dạng ATP được trình bày ở Hình 5B. ATP được tái tạo trở lại từ ADP + Pi qua hô hấp.
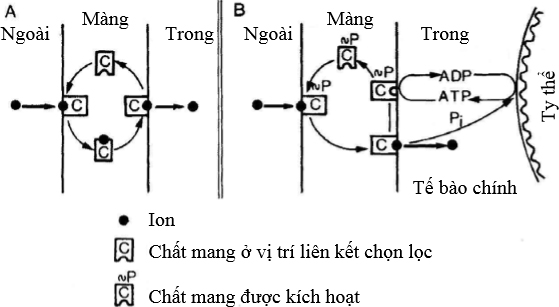
 Hình 5 Mô hình chất mang vận chuyển chất tan qua màng
Hình 5 Mô hình chất mang vận chuyển chất tan qua màng
4.2. Sự vận chuyển chủ động và thụ động: bơm điện
* Vận chuyển thụ động “downhill”: là sự khuyếch tán các chất tan ngang qua màng nhờ vào sự chênh lệch nồng độ của chất tan (vận chuyển từ nồng độ cao đến thấp), nhờ vào chất mang, khuếch tán qua các pha lipid và ngang qua các lỗ có nước (Hình 6).
* Hấp thu chủ động “uphill”: là sự vận chuyển các chất tan qua màng ngược với sự chênh lệch nồng độ (vận chuyển từ nồng độ thấp đến cao). Sự vận chuyển có liên quan tới cơ chế tiêu thụ năng lượng của “bơm” ở màng (Hình 6).
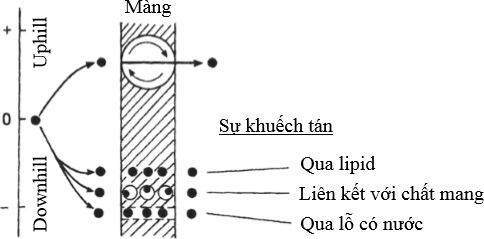
Hình 6 Mô hình vận chuyển chủ động và thụ động chất tan qua màng tế bào
5. Đặc tính của sự hấp thu ion bởi rễ
5.1. Tính chất lý hóa của các ion và sự biến dưỡng ở rễ
Trong suốt quá trình vận chuyển, chất tan từ dung dịch bên ngoài vào bên trong rễ, có tương tác giữa các chất tan (ion) và nhóm mang điện âm ở khoảng trống tự do của vách tế bào. Sự tương tác qua lại này ảnh hưởng trực tiếp đến sự hấp thu chất tan vào trong tế bào (vào tế bào chất và không bào). Một vài tính chất lý hóa của các ion và các chất tan như đường kính và hóa trị của nó, xác định tốc độ vận chuyển qua màng.
* Đường kính ion
Đối với các ion có cùng hóa trị, thường thì có tương quan nghịch giữa tốc độ hấp thu và bán kính ion, chẳng hạn khi so sánh các ion Li, Na và Kali (Bảng 3). Mặc dù Cs+ có đường kính ion nhỏ hơn, có tốc độ hấp thu nhỏ hơn ion kali. Rõ ràng là ngoài yếu tố như đường kính ion còn có những yếu tố khác có liên quan tới sự hấp thu ion của tế bào.
Bảng 3 Tương quan giữa đường kính ion và sự hấp thu cationa (Conway, 1981)
|
Cation |
Đường kính ion, nm |
Vận tốc hấp thu, µmol/g trọng lượng tươi/3 giờ |
|
Lithium |
0,38 |
2 |
|
Sodium |
0,36 |
15 |
|
Potassium |
0,33 |
26 |
|
Cesium |
0,31 |
12 |
a Cation được cung cấp ở pH 6,0 như muối bromide, 5 mM (Jacobson et al., 1960)
* Phân tử đối với sự hấp thu ion và vai trò hóa trị
Màng tế bào có cấu tạo gồm các thành phần phospholipid, sulpholipid và protein. Các thành phần nầy có mang điện tích; và các ion (chất tan) tương tác với các nhóm có mang điện tích nầy. Cường độ tương tác gia tăng theo trình tự sau:
Phân tử không điện tích < Cation+, Anion- < Cation2+, Anion2- < Cation3+, Anion3-
Trái lại, tốc độ hấp thu thường giảm theo trình tự trên. Sự gia tăng đường kính của các ion thủy hóa với hóa trị, chắc chắn là yếu tố tác động thêm cho trình tự này.
Đối với những loại chất tan không tích điện thì sự thẩm thấu ngang qua màng tế bào nhiều hơn. Thí dụ ở Hình 7 cho thấy tốc độ hấp thu của Boron giảm nhanh khi pH của dung dịch bên ngoài gia tăng. Điều này có liên quan chặt tới sự phân ly của acid boric thành anion borate khi pH gia tăng và chúng ta thấy rằng các phân tử acid boric có thể thẩm thấu qua màng nhiều hơn là các anion borate.

Hình 7 Sự hấp thu boric acid và ion borate
* Hoạt động biến dưỡng
Sự vận chuyển các ion và chất tan đòi hỏi sự tiêu hao năng lượng một cách trực tiếp và gián tiếp. Nguồn năng lượng chính là từ quá trình hố hấp trong các tế bào và mô. Các yếu tố ảnh hưởng tới quang hợp đều có ảnh hưởng tới sự tích lũy ion như:
- Oxygen. Khi tình trạng oxygen quanh rễ giảm thì sự hấp thu các ion như kali và phosphate cũng giảm theo. Đặc biệt khi oxygen có nồng độ rất thấp, trong trường hợp đất bị ngập nước, hậu quả thiếu dinh dưỡng xảy ra đã làm hạn chế sinh trưởng của cây.
- Nhiệt độ. Các phản ứng hóa học lệ thụộc nhiều vào nhiệt độ. Sự gia tăng nhiệt độ 10oC thường làm gia tăng phản ứng hóa học gấp đôi (Q10 = 2). Đối với các phản ứng enzyme thì trị số Q10 cao hơn 2. Đối với sự hấp thu của ion kali, Q10 thường cao hơn 2 trong dãy nhiệt độ sinh lý (Hình 8). Việc so sánh trị số Q10 đối với sự hấp thu ion và sự hô hấp cho thấy sự hấp thu ion lệ thụộc vào nhiệt độ nhiều hơn, đặt biệt ở nhiệt độ dưới 100C.
- Carbohydrates (nguồn năng lượng chính). Do điều kiện nguồn cung cấp carbohydrate từ lá bị giới hạn, có sự tương quan chặt giữa hàm lượng carbohydrates và sự hấp thu ion của rễ.
- Ánh sáng. Ở các tế bào quang hợp có mối tương quan chặt giữa ánh sáng, quang hợp và sự hấp thu ion. Như vậy ánh sáng ảnh hưởng trực tiếp lên sự hấp thu ion của mô lá xanh.
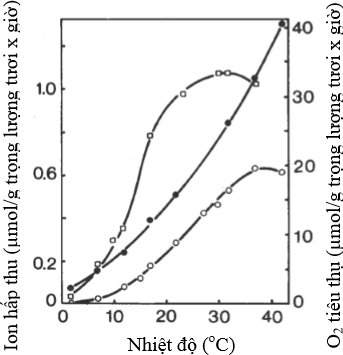
Hình 8 Ảnh hưởng nhiệt độ đến hô hấp và hấp thu ion
Có mối quan hệ chặt giữa ánh sáng, sự cung cấp carbohydrate cho rễ, sự hô hấp ở rễ và sự hấp thu ion bởi rễ, đặc biệt là khi trồng dầy, ánh sáng cung cấp cho các lá ở phía dưới bị giới hạn và các lá này là nguồn cung cấp carbohydrate chủ yếu cho rễ.
5.2. Tác động qua lại giữa các ion
* Sự cạnh tranh
Sự vận chuyển ion từ dung dịch bên ngoài vào bên trong tế bào chất đòi hỏi sự liên kết giữa ion và chất mang trên bề mặt màng tế bào chất. Vì vậy, sự cạnh tranh giữa các ion có cùng điện tích sẽ xảy ra, vì số lượng vị trí liên kết thấp hơn số lượng hay nồng độ ion cạnh tranh. Sự cạnh tranh đặc biệt xảy ra giữa các ion có các thành phần sinh lý, sinh hóa giống nhau (cùng hóa trị và đường kính ion) được trình bày ở Bảng 4.
Ở nồng độ thấp, ảnh hưởng cạnh tranh giữa Na+ hoặc Cs+ trên sự hấp thu K+ không đáng kể, còn Ca2+ kích thích sự hấp thu K+. Sự hấp thu Cs+ bị ức chế nhiều hơn khi có sự hiện diện của K+ và Rb+, điều này cho thấy cả hai cation này có ái lực với chất mang mạnh hơn là Cs+.
Sự cạnh tranh giữa các ion có hóa trị một, như giữa K+ và NH + thì khó giải thích một cách đơn giản dựa vào ái lực với chất mang. Trong khi NH + thì hoàn toàn có sự cạnh tranh với K+, nhưng ngược lại sự ức chế của K+ lên sự hấp thu NH + thì gần như không có. Để giải thích vấn đề nầy, một số tác giả cho rằng ít nhất một phần đạm ammonium được hấp thu không phải ở dạng NH +, mà là ở dạng NH thẩm thấu qua màng sau khi loại bỏ đi một proton, để lại H+ trong dung dịch bên ngoài.

Các dưỡng chất khoáng được hấp thu như các cation, ái lực của Mg2+ thủy hóa với chất mang thì đặc biệt thấp. Các ion như K+ và Ca2+ cạnh tranh với Mg2+ và vì vậy tốc độ hấp thu của Mg2+ bị giảm nhiều (Bảng 5). Sự cạnh tranh này được thấy khi bón phân nhiều kali và canxi cho cây gây ra tình trạng thiếu Mg.

Tính cạnh tranh nầy, cũng được tìm thấy giữa các anion với nhau, như sự cạnh tranh giữa Cl- và NO-, sự hấp thu NO- bị giảm bởi sự hiện diện của Cl-. Ở đất mặn, việc gia tăng nồng độ NO - làm giảm đi đáng kể sự hấp thu Cl- của cây.
* Vai trò của pH
Sự cạnh tranh giữa H+ với các cation và giữa OH- với các anion có tầm quan trọng trong dinh dưỡng khoáng của cây. Thường thì dung dịch đất ở vùng khí hậu ẩm ướt có pH < 7, nên sự cạnh tranh giữa H+ và các cation được chú ý nhiều hơn giữa OH- và anion. Ảnh hưởng của pH trong dung dịch bên ngoài đến sự hấp thu cation được trình bày ở Hình 9. Khi nồng độ ion H+ gia tăng mà không có Ca2+ sẽ làm giảm sự hấp thu K+. Ảnh hưởng này được giải thích là do sự cạnh tranh giữa H+ và K+ ở các vị trí liên kết của màng tế bào chất, trong khoảng pH từ 4 - 7. Không có Ca2+ ở pH < 4 xảy ra sự di chuyển của kali đi ra là do màng tế bào bị tổn thương, điều này cho thấy chức năng của màng sinh chất như là rào cản đối với sự khuyếch tán các ion qua màng. Sự gia tăng của Ca2+ đã làm giảm H+ sinh ra có liên quan đến sự hấp thu K+.

Hình 9 Ảnh hưởng pH đến sự hấp thu K
* Hiện tượng hiệp lực ion và vai trò của Ca2+
Hiện tượng hiệp lực ion là sự tác động ion qua lại trong suốt tiến trình hấp thu dưỡng chất. Hiện tượng hiệp lực ion có thể là kết quả sự gia tăng hoạt động biến dưỡng của rễ khi được cung cấp dưỡng chất khoáng sau một thời kỳ cạn kiệt.
Kích thích hấp thu K+ bởi Ca2+ gia tăng đồng thời giảm pH cho thấy rằng Ca2+ làm mất ảnh hưởng của ion H+ đến sự hấp thu K+. Tương tự, kích thích hấp thu K+ bởi Ca2+ giảm đi đồng thời gia tăng pH cho thấy rằng Ca2+ ức chế hấp thu K+, có lẽ đây là do sự cạnh tranh cation. Hơn nữa, bản chất của sự tác động qua lại giữa Ca2+ và K+ thay đổi tùy theo tình trạng dinh dưỡng của rễ. Ở rễ có hàm lượng K+ cao (“muối cao”), sự hấp thu K+ được kích thích do Ca2+ ở pH thấp chủ yếu là do làm giảm sự đi ra của K+. Trái lại, ở rễ có nồng độ K+ thấp (“muối thấp”), Ca2+ kích thích sự hấp thu K+ do làm gia tăng sự đi vào. Kích thích của Ca2+ cũng rõ đối với sự đi vào của Cl- và tương tự kích thích SO2- và các loại anion khác.
Do Ca2+ có thể bị tách ra khỏi vị trí liên kết của màng tế bào bởi các chất chelate hoặc có thể được trao đổi bởi H+ và các cation kim loại (như aluminum), nồng độ của Ca2+ trong dung dịch bên ngoài cần thiết để duy trì tính nguyên và tính chọn lọc của màng tế bào tùy thuộc vào pH và nồng độ của các cation cạnh tranh. Ở đất mặn, Ca2+ thực chất có thể cải thiện sự sinh trưởng cây, đặc biệt ở những cây trồng nhạy cảm với nồng độ Na+ cao trong mô.
5.3. Mối quan hệ cation - anion
Do sự hấp thu cation và anion theo qui luật khác nhau, tương tác trực tiếp giữa chúng không nhất thiết xảy ra. Tốc độ hấp thu của cation và anion khác nhau đòi hỏi có sự bù trừ điện tích cả bên trong tế bào và bên ngoài dung dịch. Mối quan hệ giữa sự hấp thu cation-anion và những thay đổi trong hàm lượng acid hữu cơ được khám phá. Sự thay đổi acid hữu cơ có liên quan tới tốc độ cố định CO2 khác nhau (sự cố định ở pha tối) trong mô rễ. Tỷ lệ hấp thu cation/anion ảnh hưởng không chỉ hàm lượng của các anion acid hữu cơ trong tế bào mà còn ảnh hưởng đến pH ở dung dịch bên ngoài. Sự hấp thu anion nhiều làm gia tăng pH bên ngoài, trái lại sự hấp thu cation nhiều thì làm giảm pH.
Có khoảng 70% nitrate (anion) hoặc ammonium (cation) được cây trồng hấp thu (Jungk, 1970). Dạng đạm cây trồng hấp thu có ảnh hưởng đến cả hàm lượng acid hữu cơ trong tế bào và pH dung dịch bên ngoài. Việc gia tăng pH do sự hấp thu nitrate là kết quả không chỉ do sự hấp thu anion nhưng cũng do sự khử nitrate trong cây, khi đó phóng thích ra đương lượng OH- theo phương trình phản ứng sau:
NO3- + 8e- + 8H+ -> NH3 + 2H2O + OH-
Nếu quá trình này xảy ra trong rễ, OH- có thể trung hòa H+ trong tế bào chất hoặc phóng thích vào trong dung dịch ngoài. Vì vậy, dạng đạm cung cấp ảnh hưởng đáng kể cả thành phần khoáng và hàm lượng acid hữu cơ của cây.
5.4. Nồng độ bên ngoài
Sự hấp thu dưỡng chất khoáng tùy thụộc vào nồng độ của dưỡng chất đó trong dung dịch bên ngoài, thí dụ như ở K+ và Na+ trong Hình 10. Sự khác nhau về hấp thu giữa K+ và Na+ cho thấy khác nhau về ái lực giữa hai ion nầy với chất mang ở màng tế bào chất của tế bào rễ. Sự hấp thu của phosphate tương tự như K+, trong khi đó sự hấp thu của Ca2+ và Mg2+ thì tương tự như Na+.
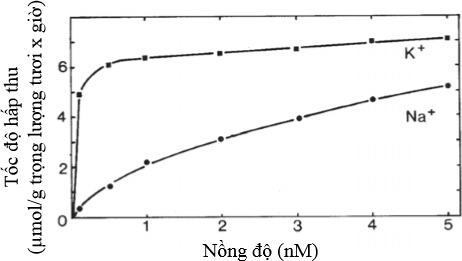
5.5. Nồng độ bên trong và tình trạng dinh dưỡng
Tương quan giữa nồng độ dưỡng chất khoáng bên ngoài và sự hấp thu còn tùy thuộc vào nồng độ của dưỡng chất khoáng ở bên trong tế bào, nghĩa là tình trạng dinh dưỡng của cây. Người ta thấy rằng ở đỉnh rễ của cây đậu, tốc độ hấp thu P tương quan chặt tới nồng độ P vô cơ trong không bào, trái lại không tương quan với nồng độ P trong tế bào chất.
Việc cung cấp P trở lại sau một thời gian thiếu P đã làm gia tăng hàm lượng P rất lớn ở chồi và cũng có thể làm ngộ độc. Những thay đổi xảy ra nhanh chóng về hàm lượng P trong dịch đất dường như không xảy ra trong đất trồng, nhưng trồng cây trong dung dịch dinh dưỡng thì yếu tố này cần được quan tâm, đặc biệt khi thay thế dung dịch dinh dưỡng mới.
6. Sự hấp thu ion dọc theo rễ
Cấu trúc và sinh lý của rễ thay đổi dọc theo trục dọc của rễ. Vận tốc hấp thu ion khác nhau ở các vùng khác nhau trên rễ. Nhìn chung, tốc độ hấp thu ion trên đơn vị chiều dài rễ giảm khi các khoảng cách càng xa chóp rễ. Cách này tương tự như sự hấp thu nước dọc theo rễ đã được Sanderson (1983) mô tả. Có 3 yếu tố chính ảnh hưởng tới sự giảm vận tốc hấp thu: (a) do sự hình thành superin ở lớp biểu bì rễ, (b) hình thành lớp vỏ trong thứ cấp ngăn cản sự vận chuyển dưỡng chất vào trung trụ, và (c) sự thoái hóa đặc biệt các tế bào phần vỏ và sự hình thành các xoang hay các khoảng trống giữa các tế bào ở phần vỏ, đôi khi có liên quan tới sự hình thành các mô khí. Cơ chế hình thành và vai trò của mô khí có liên quan tới tính thích nghi của cây trong đất bị ngập nước.
7. Sự vận chuyển qua rễ
Có hai cách vận chuyển chất tan qua phần vỏ tới trung trụ của rễ; cách thứ nhất là chất tan đi qua khoảng trống giữa các tế bào (khoảng gian bào) và khoảng trống tự do ở vách tế bào; cách thứ hai là chất tan đi từ tế bào này sang tế bào khác bằng sợi liên bào (Hình 11). Sự vận chuyển trong khoảng gian bào bị chặn lại bởi dãy Casparian ở vách của các tế bào nội bì. Dãy này bị superin hóa và bao quanh mỗi tế bào; nó hình thành rào cản giữa phần vỏ và trung trụ. Sự vận chuyển chất tan từ phần vỏ vào trong trung trụ và các chất đồng hóa vận chuyển từ trung trụ vào phần vỏ phải đi qua các tế bào nội bì. Tuy nhiên, có một vài vùng bị "rò rĩ" ở rào cản được tìm thấy nơi đỉnh sinh trưởng ngọn, ở đó có sự phân chia tế bào chưa hoàn toàn và ở vùng đáy rễ có các rễ bên đi xuyên qua nội bì. Các rễ bên phát triển từ trụ bì vào trong trung trụ. Sự vận chuyển chất tan từ bên ngoài đi vào khoảng gian bào của vùng trung trụ thì rất quan trọng, đặc biệt đối với calcium, magnesium và aluminum bởi vì sự di chuyển của các nguyên tố khoáng này bằng con đường đi qua tế bào bị giới hạn. Tầm quan trọng của mỗi con đường trong sự hấp thu dưỡng chất ở phần vỏ lệ thụộc vào nhiều yếu tố như nồng độ ion bên ngoài, sự hình thành lông rễ và sự superin hóa của các tế bào rễ.

Hình 11 Chuyển vận chất tan vào rễ xuyên qua tế bào (A) và qua những khoảng gian bào (B)
Các ion di chuyển vào trong khoảng gian bào bị hạn chế nơi vùng đáy rễ, do superin hóa của các tế bào rễ và do các chất nhầy ở vùng chóp rễ hình thành một lớp dầy ở mặt ngoài vách tế bào vùng rễ. Lớp nhầy nầy hoạt động như rào ngăn cản dòng vận chuyển thụ động của các chất tan vào bên trong rễ, do vậy các cation đa hóa trị như aluminum tích lũy lại ở lớp nhầy này.
Sự vận chuyển chất tan bằng con đường từ tế bào này sang tế bào khác nhờ cầu nối gọi là cầu liên bào, cầu nối nầy xuyên ngang qua vách tế bào mà liên kết tế bào chất với các tế bào lân cận. Helder và Boerma (1969) tìm thấy có khoảng 20.000 sợi liên bào/tế bào ở các tế bào nội bì của rễ lúa mạch còn non, các tế bào nội bì ở phần rễ già hơn thì có rất ít sợi liên bào, nhưng số lượng vẫn đủ cho phép sự vận chuyển nước và các ion đi qua các tế bào nội bì (Clarkson et al., 1971). Để đánh giá vai trò của 2 con đường vận chuyển chất tan ở rễ, thì cần quan tâm tới số lượng sợi liên bào trong các tế bào vùng rễ. Các tế bào rễ phát triển thành các lông rễ có nhiều sợi liên bào hơn các tế bào rễ bình thường.
8. Cơ chế phóng thích ion vào trong mạch gỗ
Sau khi các ion được vận chuyển bên trong tế bào vào đến trung trụ, hầu hết các ion này được phóng thích vào trong mạch gỗ. Vùng chóp rễ có một vài ống mạch metaxylem có chứa tế bào chất, nhưng hầu hết không chứa tế bào chất, vì vậy như là các khoảng gian bào. Sự phóng thích các ion và nước ở trung trục vào trong mạch gỗ cũng đồng thời có sự vận chuyển ngược lại vào trong khoảng trống tư do. Câu hỏi được đặt ra là sự phóng thích này là một quá trình thụ động (“sự rò rỉ”) hay là sự bài tiết chủ động. Sự bài tiết chủ động được ủng hộ nhiều hơn và có liên quan tới các chất ức chế biến dưỡng, đặc biệt là chất ức chế sự tổng hợp protein. Các hợp chất ức chế, trong đó cyclohexumide ức chế sự phóng thích các ion vào trong mạch gỗ nhưng không có sự tích lũy của chất nầy trong tế bào rễ. Mô hình vận chuyển ion vào trong mạch gỗ (Hình 12) cho thấy sự vận chuyển tích cực và thụ động xảy ra đồng thời để đưa ion từ dung dịch bên ngoài vào trong mạch gỗ. Nhu mô gỗ trong mô hình này đóng vai trò quan trọng cho sự bài tiết ion. Sự khám phá ra cấu trúc tương tự như tế bào vận chuyển trong các tế bào nhu mô gỗ ủng hộ quan điểm này.

Hình 12 Mô hình chuyển vận ion vào trong mạch gỗ (1) vận chuyển xuyên qua tế bào và (2) vận chuyển trong khoảng gian bào
9. Các yếu tố ảnh hưởng đến sự phóng thích ion vào trong mạch gỗ
9.1. Nồng độ bên ngoài
Khi nồng độ ion bên ngoài gia tăng dẫn tới nồng độ của ion trong nhựa mạch gỗ gia tăng. Tuy nhiên, nồng độ ion tương đối giảm khi nồng độ bên ngoài gia tăng. Sự chênh lệch nồng độ giữa dung dịch bên ngoài và nhựa mạch gỗ giảm, thậm chí có thể giảm dưới 1 trong trường hợp của calcium. Dòng khối nhựa di chuyển có nhiều hình dạng khác nhau và đạt tối đa khi nồng độ bên ngoài là 1,0 mM (Bảng 6). Khi nồng độ bên ngoài là 0,1 mM sự di chuyển nầy bị giới hạn bởi nồng độ
Bảng 6 Tương quan giữa nồng độ ion trong dung dịch bên ngoài, nồng độ ion trong nhựa và dòng khối nhựa di chuyển ion trong mạch gỗ.
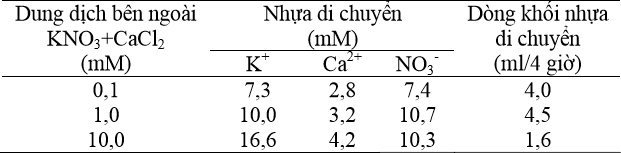
Trái lại, nồng độ bên ngoài là 10,0 mM dòng nhựa bị giới hạn do sự hiện diện của nước (nghĩa là tiềm năng nước cao trong dung dịch bên ngoài) và chênh lệch nồng độ không nhiều giữa dung dịch bên ngoài và mạch gỗ.
9.2. Nhiệt độ
Nhiệt độ rễ gia tăng ảnh hưởng đến dòng khối nhựa di chuyển nhiều hơn là ảnh hưởng đến nồng độ ion trong nhựa (Bảng 7). Nghĩa là nhiệt độ quyết định tốc độ phóng thích ion vào trong mạch gỗ và sự di chuyển của nước.

9.3. Hô hấp
Tốc độ phóng thích ion vào trong mạch gỗ tương quan chặt với hô hấp của rễ. Thiếu oxygen dòng khối nhựa di chuyển giảm mạnh, nhưng nồng độ của kali và calcium trong nhựa không giảm (Bảng 8). Trái với ảnh hưởng của nhiệt độ (Bảng 7), sự ức chế hô hấp làm chậm lại mức độ vận chuyển nhưng không ảnh hưởng đến tỷ lệ vận chuyển K+/Ca2+.

9.4. Ion đồng hành
Sự cân bằng cation-anion cần thiết cho việc phóng thích các ion vào trong mạch gỗ. Tuy nhiên, ion đồng hành dường như có ảnh hưởng đến tốc độ vận chuyển ngay cả nồng độ bên ngoài thấp. Bảng 9 cho thấy khi cung cấp KNO3 thì tốc độ di chuyển của nhựa hầu như cao gấp hai lần so với cung cấp một lượng tương đương K2SO4. Tuy nồng độ của K gần như không thay đổi, nhưng nồng độ của nitrate và sulfate trong nhựa khác nhau rất lớn. Sự khác nhau về điện tích âm trong nhựa được bù đắp lại bởi sự gia tăng nồng độ của các anion acid hữu cơ.

Khi cung cấp K2SO4, yếu tố làm hạn chế tốc độ di chuyển của nhựa có lẻ là do rễ cố gắng duy trì sự cân bằng cation-anion bằng cách tổng hợp acid hữu cơ. Điều này làm giảm tốc độ phóng thích của kali và calcium vào trong mạch gỗ và chính vì vậy vận tốc di chuyển của nhựa cũng giảm một cách tương ứng.
9.5. Tình trạng carbohydrate ở rễ
Sự phóng thích ion vào mạch gỗ có tương quan chặt với tình trạng carbohydrate của rễ. Quang kỳ có ảnh hưởng đến tình trạng carbohydrate của rễ vì vậy ảnh hưởng tới vận tốc và dòng nhựa di chuyển. Sự hấp thu và tốc độ vận chuyển kali ở rễ có hàm lượng carbohydrate cao thì lớn hơn nhiều so với rễ có hàm lượng carbohydrate thấp (Bảng 10).

Tài liệu tham khảo
Branton, D. (1969). Membrane structure. Annu. Rev. Plant physiol. 20, 209-238.
Clarkson, D. T., Robards, A. W. and Sanderson, J. (1971). The tertiary endodermis in barley roots: Fine structure in relation to radial transport of ions and water. Planta 96, 292-305.
Conway, B. E. (1981). “Ionic Hydration in Chemistry and Biophysics”. Elsevier, Amsterdam.
Helder, R. J. and Boerma, J. (1969). An electron-microscopical study of the plasmodesmata in the roots of young barley seedlings. Acta Bot. Neerl. 18, 99-107
Keller, P. and Deuel, H. (1957). Kationenaustauschkapazitat und Pektingehalt von Pflanzenwurzeln. Z. Pflanzenernaehr., Dueng., Bodenkd. 79, 119-131
Sanderson, J. (1983). Water uptake by different regions of barley root. Pathway of radial flow in relation to development of the endodermis. J. Exp. Bot. 34, 240-253.
Schimansky, C. (1981). Der Einfluss einiger Versuchsparameter auf das Fluxverhalten von 28Mg bei Gerstenkeimpflanzen in Hydrokulturversuchen. Landwirtsch. Forsch. 34, 154-165.
Singer, S. J. (1972). A fluid lipid-globular mosaic model of membrane structure. Ann. N. Y. Acad. Sci. 195, 16-23.

-
 Phác đồ hồi phục cây trồng sau ngộ độc thuốc cỏ, thuốc BVTV
Phác đồ hồi phục cây trồng sau ngộ độc thuốc cỏ, thuốc BVTV
-
 Silic – dưỡng chất quan trọng nhưng còn nhiều tranh cãi
Silic – dưỡng chất quan trọng nhưng còn nhiều tranh cãi
-
 Ứng dụng chitosan trong nông nghiệp giải pháp xanh thay thế hóa chất
Ứng dụng chitosan trong nông nghiệp giải pháp xanh thay thế hóa chất
-
 Khám Phá Bí Mật Nốt Sần Ở Cây Họ Đậu - Cơ Chế Cố Định Đạm Giúp Làm Giàu Đất
Khám Phá Bí Mật Nốt Sần Ở Cây Họ Đậu - Cơ Chế Cố Định Đạm Giúp Làm Giàu Đất
-
 Tìm hiểu các nhóm vi khuẩn nốt sần cố định đạm và tầm quan trọng trong nông nghiệp
Tìm hiểu các nhóm vi khuẩn nốt sần cố định đạm và tầm quan trọng trong nông nghiệp
-
 Sử dụng Ethephon 40% để lên màu, chín nhanh cam Canh và cam Vinh mà không rụng lá, rụng trái
Sử dụng Ethephon 40% để lên màu, chín nhanh cam Canh và cam Vinh mà không rụng lá, rụng trái

 Cây mai chiếu thủy
Cây mai chiếu thủy
 Cây đào
Cây đào
 Cây hoa hồng
Cây hoa hồng
 Cây nhãn
Cây nhãn
 COMBO COMBI 05: Biến vàng thành xanh
COMBO COMBI 05: Biến vàng thành xanh
 COMBO T-ROOT kích rễ toàn diện - phục hồi cây yếu, thúc rễ cực mạnh
COMBO T-ROOT kích rễ toàn diện - phục hồi cây yếu, thúc rễ cực mạnh
 Sử dụng kết hợp đạm Ure và Atonik đậm đặc - 'Công thức vàng' trong sử dụng phân bón
Sử dụng kết hợp đạm Ure và Atonik đậm đặc - 'Công thức vàng' trong sử dụng phân bón
 Những tác dụng và tính năng nổi bật khi sử dụng Cytokinin DA6 trên cây trồng
Những tác dụng và tính năng nổi bật khi sử dụng Cytokinin DA6 trên cây trồng
 Làm cách nào để ngăn chặn lộc, đọt non trên cây trồng?
Làm cách nào để ngăn chặn lộc, đọt non trên cây trồng?
 Hướng dẫn kích thích hoa ly nở và kìm hãm hoa cúc nở vào đúng dịp Tết
Hướng dẫn kích thích hoa ly nở và kìm hãm hoa cúc nở vào đúng dịp Tết
 Làm cách nào để cây hoa cúc phát triển nhanh và nở hoa đúng thời điểm?
Làm cách nào để cây hoa cúc phát triển nhanh và nở hoa đúng thời điểm?
 Cơ chế hút nước và dinh dưỡng của cây trồng
Cơ chế hút nước và dinh dưỡng của cây trồng
 Đất là gì? khái niệm về đất, bản chất và thành phần của đất
Đất là gì? khái niệm về đất, bản chất và thành phần của đất
 Quá trình hấp thụ và vận chuyển khoáng trong cây trồng
Quá trình hấp thụ và vận chuyển khoáng trong cây trồng
 Đất mặn và ảnh hưởng của đất mặn đến sự phát triển của cây trồng
Đất mặn và ảnh hưởng của đất mặn đến sự phát triển của cây trồng

 Trang chủ
Trang chủ
 Cây trồng phong thủy
Cây trồng phong thủy
 Cây CN dài ngày
Cây CN dài ngày
 Cây CN ngắn ngày
Cây CN ngắn ngày
 Cây lương thực
Cây lương thực
 Cây rau màu
Cây rau màu
 Cây ăn quả (trái)
Cây ăn quả (trái)
 Cây cảnh, hoa cảnh
Cây cảnh, hoa cảnh
 Cây thuốc (dược liệu)
Cây thuốc (dược liệu)
 Cây lâm nghiệp
Cây lâm nghiệp
 Cẩm nang phân bón
Cẩm nang phân bón
 Trồng trọt, chăm sóc
Trồng trọt, chăm sóc
 Khoa học nông nghiệp
Khoa học nông nghiệp
 Sức khỏe và làm đẹp
Sức khỏe và làm đẹp
